For many years, both human and animal studies correlated changes in behaviour of the young offspring with the degree of maternal stress or glucocorticoid exposure of the foetus/neonate. In the past ten years there has been overwhelming epidemiological evidence to suggest that growth retardation in utero is a very important risk factor for the development of cardiovascular and metabolic disease in adult life. More recently, it has been shown that one important, even key, determinant is the exposure of the foetus to excess glucocorticoid. Even a brief period (48 h) of dexamethasone exposure very early in pregnancy was able to programme permanently hypertensive adult sheep. Understanding how such programming works, and the underlying physiological changes that occur, provides one of the most exciting challenges in contemporary endocrinology and developmental biology.
For many years, the only determinants of non-communicable disease in the adult were seen as life-style factors and genetic predisposition. More recently, it has been suggested that the first environment, the womb, might be as crucial, if not more crucial, than either of these two factors alone. This has introduced the concept of ’programming’, whereby some change from normal, occurring at a specific, restricted time in development might have long-lasting consequences, setting in train a series of events that culminate in the adult onset of disordered function. There have been some recent reviews on this topic1–5. The current review will cover some of the same ground but will attempt to give an overview of the exciting areas that are under current investigation. Therefore, the reference list will contain a larger proportion than usual of meeting abstracts.
In fact, the concept that the uterine environment might have an important influence on the subsequent fate of the offspring is not really new, it is just that the literature in this area concentrated on factors that might influence the behaviour of the neonate/adolescent/adult. If any disturbances were recorded they were likely to be behav-ioural or psychiatric in nature. Thus, there are numerous papers in which the behaviour of the offspring is correlated with the influence of a large variety of ’stresses’ experienced by the mother. These ’stresses’ include the repeated exposure to loud, unanticipated noise (such as experienced by people living under flight paths of busy airports) and living in a country preparing for and ultimately going to war (such as the six-day Israeli war). The offspring showed intrauterine growth retardation, and/or delays in attaining motor, verbal and social skills6. Experimental animal studies were largely directed at determining how the brain had been affected by the presumed mediators – adrenal glucocorticoid hormones7.
However, about ten years ago, epidemiological studies from the UK (Ref. 8) began to highlight the association between inappropriately low birth weight and significantly increased risks of the adult onset of cardiovascular disease (which affects 20% of people over 45 years of age). This was subsequently extended to associations with the risk of developing metabolic diseases – type 2 diabetes mellitus and dyslipidaemia9,10. The ’Barker hypothesis’ is that undernutrition occurring before birth leads to both growth retardation of the foetus and to adaptations made by the foetus to cope with the lack of particular nutrients. These permanently alter physiological systems leading, ultimately, to pathological consequences1. Potential mechanisms are shown inFig. 1, and discussed in greater detail later in the review.
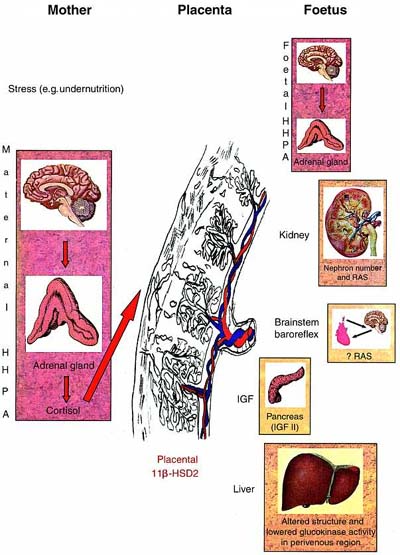 |
| Figure 1. Potential mechanisms involved in foetal programming. Stress to the mother (for example, undernutrition) may activate the maternal hippocampal–hypothalamic–pituitary–adrenal axis (HHPA), which, depending on the level of the placental corticosteroid inactivating enzyme, 11-hydroxysteroid dehydrogenase type 2 (11-HSD2), will expose the foetus to high concentrations of glucocorticoid (GC). High levels of GC in the foetus will alter the number of steroid receptors in the HHPA, affect the level of activity of other systems such as the growth hormone/insulin-like growth factor (IGF) axis, affect the replacement of foetal-type pancreaticcells with adult-type cells, and affect the renin–angiotensin system (both systemic and local) which will, ultimately, lead to pathological changes in adulthood. |
By 1996, the link between adult cardiovascular disease and low birth weight (for gestational age) had been found in 34 studies involving more than 60 000 subjects11. Subsequently, at least one study did not confirm these findings12 but another did13. Continuing studies from Barker and colleagues reported that people of birth weights 2.5 kg had, at age 50 years, higher resting pulse rates than those who had weighed 3.3 kg or more14. This was interpreted as evidence of sympathetic nerve activity being programmed in utero. In addition, this group found that even within the normal range of resting plasma cortisol concentrations, higher values occurred in the lowest birth weight group, and were related to systolic blood pressure, plasma triglyceride levels and the degree of insulin resistance15.
Experiments in guinea pigs, rats and sheep have confirmed the link between intrauterine growth retardation and adult hypertension or metabolic disease (S.L. Fielke et al., Abstract*).21–5,16,17. Regardless of the mechanism by which growth is restricted (by uterine artery ligation, by maternal anaemia, or by the restriction of either total calories or protein only), the outcome is the same. Offspring of rats fed low-protein diets during pregnancy and lactation had substantial changes in liver morphometry and greatly reduced glucokinase activity in the perivenous region18. In growth-restricted guinea pigs, the young male adults develop hypertension, show reduced skeletal muscle mass and increased adipose tissue, are glucose intolerant and develop impaired cholesterol homeostasis (K. Kind et al. and J.A. Owens et al., Abstracts..). What is most interesting is that undernutrition in the first half of pregnancy, in a long-gestation species like the sheep, leads to hypertensive lambs at three months of age16.
The evidence that glucocorticoids (GCs) might play a very important role in foetal programming of adult systemic disease came from a variety of sources. Interest in these compounds was generated by substantial evidence that GCs could directly increase blood pressure and insulin resistance in the adult19,20, and that GC exposure during pregnancy could retard foetal growth and reset the functioning of the adult hippocampal–hypothalamic–pituitary–adrenal axis (HHPA). Moreover, undernutrition was shown, in non-pregnant animals, to increase the activity of the HHPA system.121. Further studies included the use of carbenoxolone to inhibit placental inactivation of GCs, which resulted in both growth retardation and increased blood pressure in the adult offspring2,22. Maternal adrenalectomy, with only saline (0.9% NaCl) replacing drinking water, led to a reduction in birth weight but no increase in the blood pressure of the offspring at six months. The addition of carbenoxolone to the saline drunk by the adrenalectomized mothers did not produce any further reduction in birth weight or significantly change blood pressure. Metyrapone given for days 0–14 of pregnancy to rats on a low-protein diet resulted in offspring, of both sexes, with growth retardation but normal blood pressure at seven weeks. Corticosterone replacement in metyrapone-treated dams resulted in hypertensive female, but not male, offspring at six weeks2,23.
In many studies, the synthetic steroid dexamethasone was used, as it is not metabolized to the same extent by 11-hydroxysteroid dehydrogenase type 2 (11-HSD2). These studies are of interest because the question of timing, of a ’critical period’, could be addressed. Treatment of pregnant rats with dexamethasone for only the last third of gestation increased blood pressure 16 weeks after birth24. One recent remarkable finding was that treatment of pregnant sheep for only two days, at the end of the first month of pregnancy (which lasts five months) was able to programme increased blood pressure in the lambs from three months of age25. The same effect was not produced when the dexamethasone was given at the beginning of the third month. The treatment was of such short duration that there was no effect on birth weight. It fits with the data of Hawkins, who found that undernutrition only in the first half of pregnancy resulted in lambs that were hypertensive at three months16. It also shows that crucial time periods can occur early as well as late in pregnancy. This has been well accepted for the programming by androgens of neuroendocrine sexual maturation26 but not previously considered important in glucocorticoid action.
In theory, many organs or systems could be altered during foetal life with long-term consequences.
The entire RAS [renin, angiotensinogen, angiotensin-converting enzyme, receptors type 1 and 2 – (AT1, AT2)] is expressed at many sites in the foetus, including the kidney, brain and cardiovascular system27,28, and it is known to regulate blood pressure at multiple levels29. Interactions between the RAS and the insulin-mediated control of blood glucose are becoming evident30. In utero growth retardation has been shown to affect renal development particularly, with fewer nephrons being formed in the active period of nephrogenesis and higher than normal renin and angiotensin II concentrations in cord blood31,32. If fewer nephrons are available throughout postnatal life, the risk of developing renal failure, which would activate the RAS in an effort to preserve the glomerular filtration rate, could increase33. Preliminary evidence that the RAS might be involved comes from studies indicating that the blood pressure of the growth-retarded hypoxic foetal sheep seems to be maintained at normal levels by increased activity of the RAS (L.J. Edwards et al., Abstract.). Short-term (4 h) infusions of captopril, an inhibitor of the production of angiotensin II, lowered blood pressure in growth-retarded but not in normally grown foetuses. In addition, treatment of rats from undernourished mothers with captopril from weeks two to four after birth prevented the expected hypertension from developing34. In our studies, the sheep that had developed high blood pressure as a result of early treatment with dexamethasone did not show increased vascular reactivity to exogenous angiotensin II at 5–19 months of age25. However, the RAS is also involved in the setting of the baroreflex, which acts normally to set blood pressure within certain limits35. In addition, we have demonstrated that the difference in blood pressure continually increases as the animals get older (M. Dodic, unpublished data obtained at 40 months of age; seeFig. 2) and there is evidence suggesting that the baroreflex is reset (M. Dodic, unpublished). Preliminary results showing that the cardiac outputs are higher in these sheep than in the control group are in accordance with these baroreflex results.
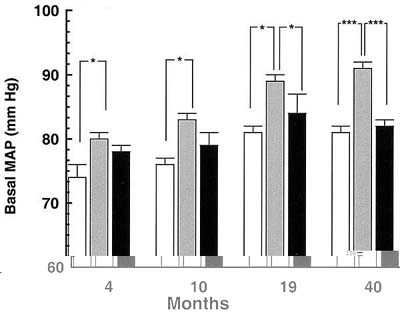 |
| Figure 2. Effect of prenatal glucocorticoid exposure on basal mean arterial pressure (MAP) of female offspring of control (n = 7) (open square); lambs exposed to dexamethasone 0.48 mg h-1 for 48 h at 27 days of gestation (n = 6) (shaded square); and lambs exposed to dexamethasone 0.48 mg h-1 for 48 h at 64 days of gestation (n = 7) (closed square) when 4, 10, 19 and 40 months old. Basal mean arterial pressure was monitored continuously by computer at 10 min intervals for three days and is represented as the mean value. Results are means + SEM. *p 0.05; ***p 0.001. |
Most evidence to date concerns changes to some component of the CNS (Ref. 36). The brainstem, hippocampus, hypothalamus and neocortex are the major areas showing changes after prenatal stress or excess glucocorticoid exposure. Rats, exposed to dexamethasone on days 17–19 of pregnancy, had reduced amounts of norepinephrine in the neocortex and the hippocampus, at 14 weeks post-partum, and altered turnover of serotonin in the hypothalamus, hippocampus and neocortex, which was detectable from three weeks of age.137. These changes in neurotransmitter content and/or metabolism were reflected in behavioural changes (hyperexcitability) and higher plasma corticosterone levels when first exposed to a novel environment. Vervet monkeys, treated with a single dose of dexamethasone at 80% of gestation, were shown by magnetic resonance imaging to have a 20–30% reduction in hippocampal volume at 20 months of age.138. These monkeys also had consistently higher plasma cortisol values (morning, afternoon and post-stress) than did vehicle-treated controls. One popular theory suggests that the ’set’ of the HHPA axis may be altered by changes in the numbers and/or affinity of glucocorticoid receptors mediating normal feedback by some ’programming’ event. In adult life, the organism is then exposed to higher than normal endogenous levels of GC. Prolonged exposure to excess GC can produce increases in blood pressure and predispose to type 2 diabetes.220. The most recent experiments show that in the foetus of the ewe undernourished in the first half of gestation the responses of the HHPA to hypoxia or exogenous corticotrophin-releasing hormone are diminished in utero, but exaggerated some time after birth.416,39.
It is now known that thecells of the foetus undergo apoptosis at some crucial stage (28–32 weeks of gestation in the human; postnatal days 7–14 in the rat) and are replaced by adultcells40,41. The hormonal system of the insulin-like growth factors is involved in this process42.
Although dexamethasone has been used widely as a ’glucocorticoid’, it may not exert all of its effects by an action on the classic GC receptor. Many years ago it was found that the predominant high-affinity binding sites for dexamethasone in bovine tissues were not classic GC receptors43. Recently, a pregnane X receptor (PXR) has been described that does not bind to naturally occurring GCs (cortisol, corticosterone) but does bind dexamethasone as well as progesterone, pregnenolone and some of their derivatives. This PXR binds as a heterodimer with the 9-cis retinoic acid receptor (RXR) to a hormone response element composed of two half-sites, organized as a direct repeat44. Potently active neurosteroids, such as allopregnanolone and tetrahydrodeoxycorticosterone, are produced by the actions of 5reductases, particularly type 2, and 3hydroxysteroid dehydrogenases (3HSDs)45. The gene for 5reductase 2 is expressed transiently in the rat brain in late gestation and early postnatal life46. The 3HSDs are inducible by dexamethasone working on elements unrelated to classic GC response elements47. Neurosteroids can affect many aspects of brain differentiation subserving endocrine and behavioural functions by interactions with theaminobutyric acid receptor, the nicotinic acetylcholine receptor, the N-methyl-d-aspartate receptor, and possibly membrane steroid receptors45,48. Neurosteroids, such as allopregnanolone, are implicated in baroreceptor reset49. These are some exciting new areas that merit further investigation.
The survival rate for preterm babies (24–26 weeks of gestation) has improved over the past 20 years, largely owing to the introduction of antenatal steroid treatment of the mother and the use of surfactant postnatally. Repeated courses of GC treatment have been used when delivery does not occur immediately. Doubts have been raised as to whether repeated courses have the potential to cause unwanted side-effects50,51. Studies in sheep have shown that repeated injections of GC can cause a delay in myelination of the optic nerve axons, and raise the question of potential damage to the CNS (Ref. 52). Some behavioural changes have been recorded in children at risk of congenital adrenal hyperplasia, owing to 21-hydroxylase deficiency, who were treated in utero with long-term low doses of GC (Ref. 53). Even though prenatal GC treatment is effective in eliminating virilization of females affected by congenital adrenal hyperplasia54, potentially, only one in eight treated foetuses benefit from the treatment55. In view of the material covered in this review, it would seem prudent to limit foetal exposure to exogenous GC to the minimum.
This review underlines the importance of the well-fed and relaxed mother (Fig. 3), but also attempts to address several very important questions for future research.
 |
| Figure 3. Cartoons depicting the baby’s need for a well-fed, relaxed mother. |
How much stress in a pregnant woman is enough to put the foetus at risk of adult cardiovascular or metabolic disease? There have been few and conflicting studies linking perceived psychosocial stress with measurement of hormonal levels56,57. What are the critical periods – is there more than one?
Is it only glucocorticoid exposure that is potentially undesirable? Can excess mineralocorticoid exposure programme undesirable effects? This has been suggested, as patients inheriting glucocorticoid-suppressible hyperaldosteronism from their mothers were found to have higher levels of plasma aldosterone and blood pressure than those inheriting the disease from their fathers58. Presumably, these people may have had chronic exposure to high aldosterone concentrations during intrauterine life. Recently, it has been found that there is a strong association between the degree of visceral fat and increased plasma aldosterone concentrations59. Increased visceral fat is one component of syndrome X for which the risk is increased more than tenfold in low birthweight babies9. Chronic exposure to excess aldosterone can cause hypertension60. Aldosterone, as well as GC, can alter the hippocampal expression of both mineralocorticoid and glucocorticoid receptor mRNAs (Ref. 61).
What are the mechanisms by which the pathological changes are induced? Is there a good marker by which one can determine the most vulnerable babies? Is there an intervention that we can use in the low birthweight baby that can reduce the risk of developing high blood pressure and/or metabolic disease 40 years later?
The work of the authors was supported by a block grant (983001) to the Howard Florey Institute from NH MRC, Australia. The authors wish to thank Dr Julie Owens and Professor Mark Hanson for provision of relevant abstracts, and Johanna Coghlan for the artwork.
[1]Barker D.J.P. (1997)Fetal nutrition and cardiovascular disease in later life.
Br. Med. Bull., 53:96-108. [MEDLINE] [Cited by]
[2] Seckl J.R. (1997) Glucocorticoids, fetoplacental 11-hydroxysteroid dehydrogenase type 2, and the early life origins of adult disease.
Steroids, 62:89-94. [ScienceDirect] [MEDLINE] [Cited by]
[3] Langley-Evans S.C. (1997) Intrauterine programming of hypertension by glucocorticoids.
Life Sci., 60:1213-1221. [ScienceDirect] [MEDLINE] [Cited by]
[4] Robinson J.S., Owens J.A. and McMillen I.C. (1997) Back to the womb.
Today’s Life Science, 9:18-25. [Cited by]
[5] Gluckman P.D. and Harding J.E. (1997) Fetal growth retardation: underlying endocrine mechanisms and postnatal consequences.
Acta Paediatr. Suppl., 442:69-72. [Cited by]
[6] Weinstock M., Fride E. and Hertzberg R. (1988) Prenatal stress effects on functional development of the offspring.
Prog. Brain Res., 73:319-330. [MEDLINE] [Cited by]
[7] de Kloet E.R. and Joels M. (1996) Corticosteroid hormones in neuroprotection and brain damage.
Curr. Opin. Endocrinol. Diabetes, 3:184-192. [Full text] [Cited by]
[8] Barker D.J.P. et al. (1989) Growth in utero, blood pressure in childhood and adult life, and mortality from cardiovascular disease.
Br. Med. J., 298:564-567. [Full text] [Cited by]
[9] Barker D.J.P. et al. (1993) Type 2 (non-insulin-dependent) diabetes mellitus, hypertension and hyperlipidaemia (syndrome X): relation to fetal growth.
Diabetologia, 36:62-67. [MEDLINE] [Cited by]
[10] Ravelli A.C.J. (1998) Glucose tolerance in adults after prenatal exposure to famine.
Lancet, 351:173-177. [ScienceDirect] [MEDLINE] [Cited by]
[11] Law C.M. and Shiell A.W. (1996) Is blood pressure inversely related to birth weight? The strength of evidence from systematic review of the literature.
J. Hypertens., 14:935-941. [MEDLINE] [Cited by]
[12] Stanner S.A. et al. (1997) Does malnutrition in utero determine diabetes and coronary heart disease in adulthood? Results from the Leningrad siege study, a cross sectional study.
Br. Med. J., 315:1342-1349. [Full text] [Cited by]
[13] Rich-Edwards J.W. et al. (1997) Birth weight and risk of cardiovascular disease in a cohort of women followed up since 1976.
Br. Med. J., 315:396-400. [Full text] [Cited by]
[14] Phillips D.I.W. and Barker D.J.P. (1997) Association between low birthweight and high resting pulse in adult life: is the sympathetic nervous system involved in programming of the insulin resistance syndrome?
Diabetes Med., 14:673-677.
[15] Phillips D.I.W. et al. (1998) Elevated plasma cortisol concentrations: a link between birth weight and the insulin resistance syndrome?
J. Clin. Endocrinol. Metab., 83:757-760. [MEDLINE] [Cited by]
[16] Hawkins P. et al. (1997) Cardiovascular development in late gestation fetal sheep and young lambs following modest maternal nutrient restriction in early gestation.
J. Physiol., 505P:18. (abstr.) [Cited by]
[17] Holemans K., van Bree R., Verhaeghe J., Meurrens K. and van Assche A.F. (1997) Maternal semistarvation and streptozotocin-diabetes in rats have different effects on the in vivo glucose uptake by peripheral tissues in their female adult offspring.
J. Nutr., 127:1371-1376. [Full text] [MEDLINE] [Cited by]
[18] Burns S.P. et al. (1997) Gluconeogenesis, glucose handling, and structural changes in livers of adult offspring of rats partially deprived of protein during pregnancy and lactation.
J. Clin. Invest., 100:1768-1774. [Full text] [MEDLINE] [Cited by]
[19] Whitworth J.A., Brown M.A., Kelly J.J. and Williamson P.M. (1995) Mechanisms of cortisol-induced hypertension in humans.
Steroids, 60:76-80. [MEDLINE] [Cited by]
[20] Walker B.R. et al. (1998) Increased glucocorticoid activity in men with cardiovascular risk factors.
Hypertension, 31:891-895. [MEDLINE] [Cited by]
[21] Jacobson L., Zurakowski D. and Majzoub J.A. (1997) Protein malnutrition increases plasma adrenocorticotrophin and anterior pituitary proopiomelanocortin messenger ribonucleic acid in the rat.
Endocrinology, 138:1048-1057. [MEDLINE] [Cited by]
[22] Langley-Evans S.C. (1997) Maternal carbenoxolone treatment lowers birthweight and induces hypertension in the offspring of rats fed a protein-replete diet.
Clin. Sci., 93:423-429. [Cited by]
[23] Langley-Evans S.C. (1997) Hypertension induced by fetal exposure to a maternal low-protein diet, in the rat, is prevented by pharmacological blockade of maternal glucocorticoid synthesis.
J. Hypertens., 15:537-544. [MEDLINE] [Cited by]
[24] Levitt N.S., Linsay R.S., Holmes M.C. and Seckl J.R. (1996) Dexamethasone in the last week of pregnancy attenuates hippocampal glucocorticoid receptor gene expression and elevates blood pressure in the adult offspring in the rat.
Neuroendocrinology, 64:412-418. [Full text] [MEDLINE] [Cited by]
[25] Dodic M., May C.N., Wintour E.M. and Coghlan J.P. (1998) An early prenatal exposure to excess glucocorticoid leads to hypertensive offspring in sheep.
Clin. Sci., 94:149-155. [Cited by]
[26] Kosut S.S., Wood R.I., Herbosa-Encarnacion C. and Foster D.L. (1997) Prenatal androgens time neuroendocrine puberty in the sheep: effect of testosterone dose.
Endocrinology, 138:1072-1077. [MEDLINE] [Cited by]
[27] Wintour E.M. (1997) The renin– angiotensin system and the development of the kidney.
Trends Endocrinol. Metab., 8:199-207. [ScienceDirect] [Cited by]
[28] Schutz S., LeMoullec J.M., Corvol P. and Gase J.M. (1996) Early expression of all the components of the renin–angiotensin-system in human development.
Am. J. Pathol., 149:2067-2079. [MEDLINE] [Cited by]
[29] Reid I.A. (1992) Interactions between Ang II, sympathetic nervous system, and baroreceptor reflexes in regulation of blood pressure.
Am. J. Physiol., 262:E763-E778. [MEDLINE] [Cited by]
[30] Tait J.F. and Tait S.A.S. (1997) Insulin, the renin–angiotensin–aldosterone system and blood pressure.
Perspect. Biol. Med., 40:246-259. [MEDLINE] [Cited by]
[31] Kingdom J.C.P., McQueen J., Connell J.M.C. and Whittle M.J. (1993) Fetal angiotensin II levels and vascular (type I) angiotensin receptors in pregnancies complicated by intrauterine growth retardation.
Br. J. Obstet. Gynaecol., 100:476-482. [MEDLINE] [Cited by]
[32] Konje J.C., Bell S.C., Morton J.J., de Chazal R. and Taylor D.J. (1996) Human fetal kidney morphometry during gestation and relationship between weight, kidney morphometry and plasma active renin concentration at birth.
Clin. Sci., 91:169-175. [Cited by]
[33] MacKenzie H.S., Lawler E.V. and Brenner B.M. (1996) Pathogenesis and pathophysiology of essential hypertension. Congenital oligonephropathy: the fetal flaw in essential hypertension?
Kidney Int., 49:S30-S34. [Cited by]
[34] Sherman R.C. and Langley-Evans S.C. (1998) Early administration of angiotensin-converting enzyme inhibitor captopril, prevents the development of hypertension programmed by intrauterine exposure to a maternal low-protein diet in the rat.
Clin. Sci., 94:373-381. [Cited by]
[35] dos Santos C.M., Moreira E.D., Krieger E.M. and Michelini L.C. (1998) Chronic AT1 receptor blockade alters aortic nerve activity in hypertension.
Hypertension, 31:973-977. [MEDLINE] [Cited by]
[36] Meaney M.J. et al. (1994) Early environmental programming of hypothalamic–pituitary–adrenal responses to stress.
Semin. Neurosci., 6:247-259. [Cited by]
[37] Muneoka K. et al. (1997) Prenatal dexamethasone exposure alters brain monoamine metabolism and adrenocortical response in rat offspring.
Am. J. Physiol., 273:R1669-R1675 . [MEDLINE] [Cited by]
[38] Uno H. et al. (1994) Neurotoxicity of glucocorticoids in the primate brain.
Horm. Behav., 28:336-348. [MEDLINE] [Cited by]
[39] Hawkins P. et al. (1998) Effect of maternal nutrient restriction in early gestation on hypothalamic pituitary adrenal axis responses during acute hypoxaemia in late gestation fetal sheep.
J. Physiol., 507P:50. (abstr.) [Cited by]
[40] Scaglia L., Cahill C.J., Finegood D.T. and Bonner-Weir S. (1997) Apoptosis participates in the remodeling of the endocrine pancreas in the neonatal rat.
Endocrinology, 138:1736-1741. [MEDLINE] [Cited by]
[41] Tornehave D. and Larsson L.I. (1997) Presence of Bcl-x1 during development of the human fetal and rat neonatal endocrine pancreas; correlation to programmed cell death.
Exp. Clin. Endocrinol. Diabetes, 105:A27. (abstr.) [Cited by]
[42] Petrick J., Arany E., McDonald T.J. and Hill D.J. (1998) Apoptosis in the pancreatic islets of the neonatal rat is associated with reduced expression of insulin-like growth factor II that may act as a survival factor.
Endocrinology, 139:2994-3004. [MEDLINE] [Cited by]
[43] Barlow J.W. et al. (1979) Predominant high affinity binding of [3H]-dexamethasone in bovine tissues is not to classical glucocorticoid receptors.
Endocrinology, 105:827-834 . [MEDLINE] [Cited by]
[44] Kliewer S.A. et al. (1998) An orphan nuclear receptor activated by pregnanes defines a novel steroid signaling pathway.
Cell, 92:73-82 . [Full text] [MEDLINE] [Cited by]
[45] Baulieu E.E. (1997) Neurosteroids: of the nervous system, by the nervous system, for the nervous system.
Rec. Prog. Horm. Res., 52:1-32.
[46] Poletti A., Negri-Cesi P., Rabuffetti M., Colciago A., Celotti F. and Martini L. (1998) Transient expression of the 5-reductase type 2 isozyme in the rat brain in late fetal and early postnatal life.
Endocrinology, 139:2171-2178. [MEDLINE] [Cited by]
[47] Huss J.M., Wang S.I., Astrom A., McQuiddy P. and Kasper C.B. (1996) Dexamethasone responsiveness of a major glucocorticoid-inducible CYP3A gene is mediated by elements unrelated to a glucocorticoid receptor binding motif.
Proc. Natl. Acad. Sci. U. S. A., 93:4666-4670. [MEDLINE] [Cited by]
[48] Wehling M. (1997) Specific nongenomic actions of steroid hormones.
Annu. Rev. Physiol., 59:365-393. [Full text] [MEDLINE] [Cited by]
[49] Laiprasert J.D., Rogers R.C. and Heesch C.M. (1998) Neurosteroid modulation of arterial baroreflex-sensitive neurons in rat rostral ventrolateral medulla.
Am. J. Physiol., 274:R903-R911. [MEDLINE] [Cited by]
[50] Seckl J.R. and Miller W.L. (1997) How safe is long-term prenatal glucocorticoid treatment?
J Am. Med. Assoc., 277:1077-1079. [MEDLINE] [Cited by]
[51] Quinlivan J.A., Evans S.F., Dunlop S.A., Beazley L.D. and Newnham J.P. (1998) Use of corticosteroids by Australian obstetricians – a survey of clinical practice.
Aust. New Zealand J. Obstet. Gynaecol., 38:1-7. [MEDLINE] [Cited by]
[52] Dunlop S.A., Archer M.A., Quinlivan J.A., Beazley L.D. and Newnham J.P. (1997) Repeated prenatal corticosteroids delay myelination in the ovine central nervous system.
J. Matern. Fetal Med., 6:309-313. [MEDLINE] [Cited by]
[53] Trautman P.D., Meyer-Bahlburg H.F.L., Postelnek J. and New M.I. (1995) Effects of early prenatal dexamethasone on the cognitive and behavioural development of young children: results of a pilot study.
Psychoneuroendocrinology, 29:439-449. [ScienceDirect] [Cited by]
[54] Forest M.G., Morel Y. and David M. (1998) Prenatal treatment of congenital adrenal hyperplasia.
Trends Endocrinol. Metab., 9:284-289. [Full text] [Cited by]
[55] Miller L.M. (1998) Prenatal treatment of congenital adrenal hyperplasia: a promising experimental therapy of unproven safety.
Trends Endocrinol. Metab., 9:290-293. [Full text] [Cited by]
[56] Dorn D.L., Suman E.J. and Petersen A.C. (1993) Cortisol reactivity and anxiety and depression in pregnant adolescents: a longitudinal per
{kind=link}
{kind=link}